
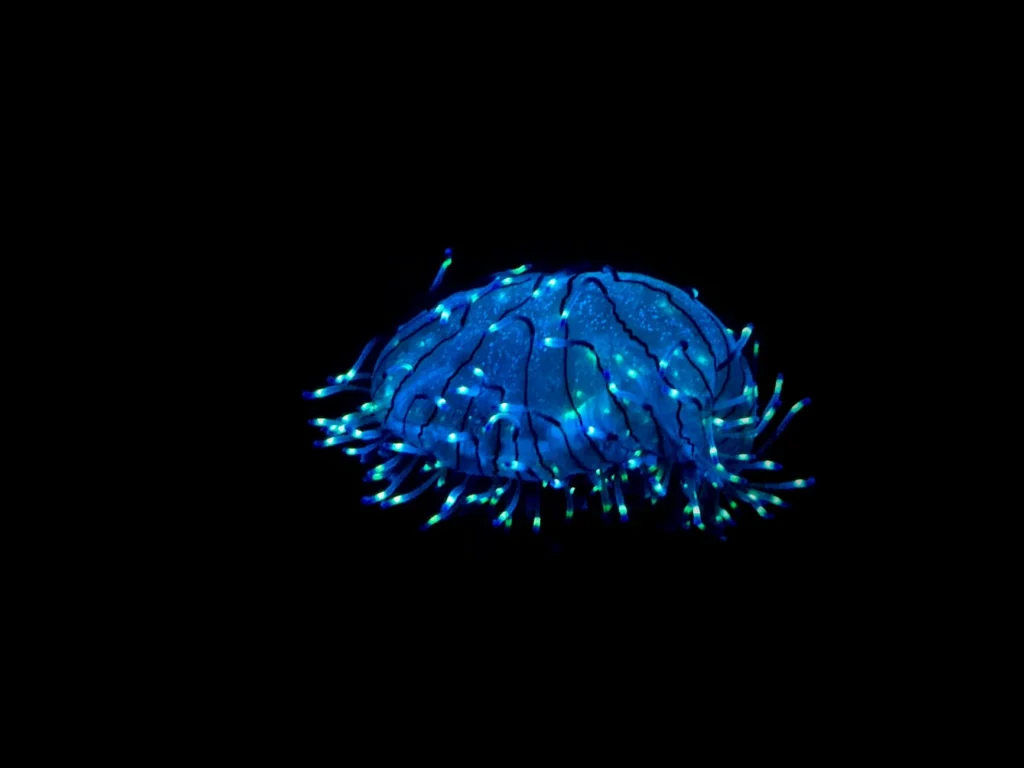
In den pechschwarzen Tiefen des Ozeans, wo kein Sonnenlicht je eindringt, besitzen schätzungsweise 75 % der Tiefseebewohner irgendeine Form der Lichtproduktion[s]. Biolumineszenz bei Tieren, die biochemische Erzeugung kalten Lichts, hat sich mehr als 50-mal unabhängig im Stammbaum des Lebens entwickelt[s]. Etwa 3.500 Arten sind dafür bekannt, auf diese Weise Licht zu erzeugen, wobei viele weitere wahrscheinlich noch unentdeckt sind[s]. Das Phänomen reicht in der Meeresumwelt rund 540 Millionen Jahre zurück und ist damit eine uralte marine Anpassung[s].
Wie Biolumineszenz bei Tieren funktioniert
Anders als eine Glühlampe, die Licht durch Wärme erzeugt, produziert Biolumineszenz bei Tieren Licht allein durch Chemie. Der Prozess umfasst zwei zentrale Akteure: ein lichtemittierendes Molekül namens Luciferin und ein Enzym namens Luciferase, das die Reaktion auslöst[s]. Wenn Luciferin in Gegenwart von Luciferase mit Sauerstoff reagiert, gibt es Energie als sichtbares Licht statt als Wärme ab.
Diese Effizienz ist bemerkenswert. Für Glühwürmchen-Luciferase wurde eine Quantenausbeute von rund 88 % gemeldet, was sie ungewöhnlich effizient im Umwandeln von Reaktionsenergie in Licht mit geringem Wärmeverlust macht[s]. Eine Glühlampe hingegen verschwendet den Großteil ihrer Energie als Wärme.
Tiere erzeugen Biolumineszenzlicht auf drei Wegen[s]: durch spezialisierte Zellen in ihrem Körper (intrazellulär), durch Ausscheidung von Chemikalien, die außerhalb ihres Körpers reagieren (extrazellulär), oder durch Beherbergung biolumineszenter Bakterien in speziellen Organen (symbiotisch). Tiefseefische nutzen häufig bakterielle Symbionten, während Glühwürmchen und Quallen Licht in ihren eigenen Zellen erzeugen.
Warum Tiere Licht erzeugen
Biolumineszente Organismen nutzen ihr Licht zur Räuberablenkung, zur Anlockung von Beute und zur Kommunikation innerhalb ihrer eigenen Art[s]. Glühwürmchen tauschen beim Balzen Lichtsignale aus, wobei jede Art ein einzigartiges Zeitmuster verwendet, um Verwechslungen zu vermeiden. Einige Weibchen der Gattung Photuris nutzen dieses System aus, indem sie die Blinkmuster anderer Arten imitieren, um Männchen anzulocken, die sie dann töten und fressen[s].
In der Tiefsee dient Biolumineszenz bei Tieren als Tarnung durch Gegenbeleuchtung: Fische und Tintenfische strahlen Licht nach unten, um dem schwachen Schimmer von oben zu entsprechen, und machen sich so für Räuber unterhalb unsichtbar[s]. Andere nutzen Licht als „Einbruchsalarm“ und blinken bei Störung auf, um größere Räuber anzulocken, die den Angreifer fressen könnten[s].
Farbwahl
Viele Meeresarten erzeugen blaues oder grünes Licht, das Meerwasser effizient durchdringt, während Landorganismen häufig gelbe oder grüne Töne produzieren[s]. Tiefseedrachenfische bilden eine Ausnahme: Sie emittieren rotes Licht, das nur für andere Drachenfische wahrnehmbar ist[s].
Medizinische Anwendungen
Forscher haben Biolumineszenz bei Tieren für die Krebsforschung nutzbar gemacht. Wissenschaftler an der Keck School of Medicine der USC entwickelten den Matador-Assay unter Verwendung von Luciferasen aus Tiefseekrebstieren und Garnelen[s]. Der Test erkennt den Tod von Krebszellen in nur 30 Minuten[s]. Das Labor hat mehr als 75 Krebszelllinien entwickelt, die marine Luciferasen exprimieren, um zelluläre Immuntherapien voranzutreiben, darunter CAR-T-Zellen der nächsten Generation[s].
Biolumineszenzbasierte photodynamische Therapie (BL-PDT) wird als weitere Krebsbehandlungsanwendung untersucht. Durch den Einsatz interner biolumineszenter Lichtquellen statt externer Laser wollen Forscher Photosensibilisatoren innerhalb von Tumormikroumgebungen aktivieren und die Gewebedurchdringungsgrenzen herkömmlicher lichtbasierter Therapien überwinden[s].
Die Biochemie der Biolumineszenz bei Tieren
Biolumineszenz bei Tieren folgt trotz ihrer über Dutzende Male unabhängigen Entwicklung einer wiederkehrenden chemischen Logik. Die Reaktion umfasst gewöhnlich die Oxidation eines Substrats (Luciferin) durch ein Enzym (Luciferase), die häufig über ein energiereiches Peroxid-Intermediat verläuft, das zu einem angeregten Produkt (Oxyluciferin) zerfällt[s]. Dieser angeregte Zustand kehrt durch Emission eines Photons in den Grundzustand zurück.
Über 40 verschiedene Biolumineszenzwege wurden identifiziert, jeder mit spezifischen Luciferinen und Luciferasen[s]. Die wiederholte Konvergenz auf Oxidation und Decarboxylierung zeigt, dass viele unabhängig entwickelte Systeme zu ähnlicher lichtproduzierender Chemie gelangt sind[s].
D-Luciferin-System (Käfer)
Glühwürmchen-Luciferase (ungefähr 60 kDa) katalysiert die ATP-abhängige Oxidation von D-Luciferin[s]. Die Reaktion benötigt Mg2+ als Kofaktor. D-Luciferin wird adenyliert und bildet einen Luciferase-Luciferin-AMP-Komplex, der mit molekularem Sauerstoff reagiert. Das resultierende angeregte Oxyluciferin emittiert beim Übergang in den Grundzustand gelbgrünes Licht (Maximum bei ~560 nm) und setzt CO2 und AMP als Nebenprodukte frei.
Für Glühwürmchen-Luciferase wurde eine Quantenausbeute von 88 % (±25 %) gemeldet[s], außergewöhnlich unter biolumineszenten Systemen. Diese Effizienz resultiert aus einer optimierten Enzymarchitektur, die nicht-strahlende Zerfallswege minimiert.
Coelenterazin-System (Meeresorganismen)
Coelenterazin-abhängige Luciferasen, darunter Renilla (36 kDa) und Gaussia (20 kDa) sowie das verwandte NanoLuc-System (19 kDa), benötigen kein ATP und sind für die Lichtproduktion auf Sauerstoff angewiesen[s]. Die Oxidation verläuft über ein Dioxetanon-Intermediat, mit Emissionsmaxima zwischen 450 und 500 nm (blaugrün). Gaussia-Luciferase, von Ruderfußkrebsen sekretiert, zeigt außergewöhnliche Thermostabilität und Katalyserate, was sie für Reporteranwendungen wertvoll macht.
Dinoflagellaten-System
Die Dinoflagellaten-Reaktion ist trotz jahrzehntelanger Forschung noch nicht vollständig verstanden. Die Lichtproduktion in diesen einzelligen Planktonorganismen findet in Organellen namens Szintillons statt. Mechanische Stimulation löst eine Signalkaskade aus, an der G-Proteine und TRP-Ionenkanäle beteiligt sind, was zu einem Protoneneinfluss führt, der den pH-Wert der Szintillons unter 6 senkt[s]. Die pH-Änderung setzt Luciferin aus seinem Bindungsprotein frei und ermöglicht die Oxidation durch Luciferase. Der gesamte Prozess, von der Stimulation bis zum Aufblitzen, dauert etwa 15 Millisekunden[s].
Evolutionäre Ursprünge
Eine Hypothese besagt, dass Biolumineszenz bei Tieren als Mechanismus zur Entgiftung reaktiver Sauerstoffspezies entstand, als die Erdatmosphäre sauerstoffreich wurde[s]. Frühe Organismen, die Sauerstoff nicht produktiv verarbeiten konnten, entwickelten Oxidationsreaktionen, die ihn abbauten, mit Lichtproduktion als Nebeneffekt. Die natürliche Selektion nutzte diese Chemie anschließend für Kommunikation, Verteidigung und Jagd.
Bei Metridinidae-Ruderfußkrebsen wird die Biolumineszenzfähigkeit durch Luciferase-Genduplikation begünstigt[s]. Studien an Metridia lucens zeigen unerwartet hohe genetische Vielfalt innerhalb von Luciferase-Genfamilien, mit Hinweisen auf reinigende Selektion, die funktionale Sequenzen erhält. Dieses Muster deutet auf starken Selektionsdruck auf biolumineszenzrelevante Merkmale im Verlauf der Evolutionsgeschichte hin.
Forschungs- und medizinische Anwendungen
Ruderfußkrebs-Luciferasen können gegenüber Glühwürmchen-Luciferase für zellbiologische Anwendungen Vorteile bieten: ATP-Unabhängigkeit, hohe Sekretierbarkeit, hohe enzymatische Aktivität unter vergleichbaren experimentellen Bedingungen und Stabilität unter physiologischen Bedingungen[s]. Der Matador-Assay, entwickelt mit Luciferasen aus Tiefseekrebstieren, erreicht Einzelzellempfindlichkeit beim Nachweis von Krebszellentod[s]. Forscher haben dieses System genutzt, um mehr als 75 Krebszelllinien zum Testen zellulärer Immuntherapien und CAR-T-Zellen der nächsten Generation zu entwickeln[s].
Biolumineszenz-Resonanzenergietransfer (BRET) ermöglicht nicht-strahlenden Energietransfer von Luciferase-Donoren zu fluoreszierenden Akzeptoren innerhalb einer Nähe von etwa 10 nm[s]. Diese Eigenschaft bildet die Grundlage der biolumineszenzbasierten photodynamischen Therapie (BL-PDT), die erforscht wird, um Photosensibilisatoren aus der Tumormikroumgebung heraus zu aktivieren[s] und die Gewebedurchdringungsgrenzen zu überwinden, die herkömmliche lichtbasierte Therapien einschränken.



